
- Silver-Fagan Alex
- AA.VV.
- Agostino Samuel
- Albini Ester
- Allegri Sara
- Altman Peggy
- Andrea Neyroz
- Antonucci Lauren A.
- Ardigò Silvia
- Arndt Klaus
- Austin Dan
- Barbi Moreno
- Barbieri Davide
- Bargossi Alberto Mario
- Bazzani Boris
- Bean Anita
- Benson Roy
- Bertuccioli Alexander
- Bianco Antonino
- Bordoni Bruno
- Bradley John
- Brandon Leigh
- Brescia Teodoro
- Broglia Roberta
- Broussal-Derval Aurélien
- Brown Jason
- Bruscia Guido
- Burt Phil
- Cánovas Linares Ricardo
- Calle Flauto
- Caloro Rocco
- Camporese Alessandro
- Carone Deborah
- Casadei Iacopo
- Caserta Roberto
- Cassarino Salvatore Antonio
- Cereda Ferdinando
- Ceriani Marco
- Cianti Giovanni
- Clifford Ross
- Confalonieri Francesco
- Connolly Declan
- Corno Claudio
- Cotrone Irene
- Coulson Morc
- Cuni Federica
- Dameli Massimo
- Daniels Jack
- De Bartolomeo Donato
- Di Monte Marco
- Diamantini Simone
- Dieguez Papì Julio
- Dragoni Graziella
- Ellsworth Abby
- Facchinetti Paolo
- Fagioli Fabrizio
- Faverzani Alfredo
- Federico Fignagnani
- Fiorin Daniele
- Fumarola Martino
- Furia Fabrizia
- Furlan Andrea
- Galloway Jeff
- Goater Julian
- Gollin Massimiliano
- Gollinucci Emanuele
- Gordon-Mallin Erica
- Graci Marco
- Guzman Daniel
- Guzman Ruben
- Hahn Michael
- Hedrick Allen
- Herdman Alan
- Herman Ellie
- Hilditch Graeme
- Hopker James e Jobson Simon
- Horschig Aaron
- Iannucci Alessandro
- Iogna Michele
- Iovieno Luca
- Jarmey Chris
- Jurasin Alexandra
- Kalym Ashley
- Kan Mark
- Keane Brian
- Kemmler Jürgen
- Koch Urs Manfred
- Kolbing Alexander
- Lai Federico
- Laurita Jennifer
- Liebman Hollis Lance
- Lo Bianco Simonetta
- Lolletti Luca
- Madonia Alessandro
- Magi Simone
- Mann Bryan
- Manocchia Pat
- Marchetti Mauro
- Marco Mazzesi
- Marongiu Gianpiero
- Massa Stefano
- Matthews Michael
- Maurone Stefano
- McCabe Delia
- McGuigan Mike
- Melvin Don
- Memmo Fabio
- MieBner Wolfgang
- Morán Esquerdo Óscar
- Morc Coulson
- Neri Marco
- Neumann Hannes
- Nottingham Suzanne
- Paoli Antonio
- Perticari Francesco
- Phelan Thomas W.
- Posabella Giovanni
- Pozzi Andrea
- Purcell Lisa
- Ragnar Rémi
- Ramsay Craig
- Ressa Maurizio
- Roca Stefano
- Schoenfeld Brad
- Scholl Peter
- Seeger Fabian
- Seijas Guillermo
- Senati Silvia
- Simone Fabrizio
- Sonthana Kevin
- Spilio Katerina
- Squillante Antonio
- Stecchi Alfredo
- Striano Philip
- Tarullo Roberto
- Timón Vicky
- Todea Noe
- Torri Barbara
- Tozzi Nicoletta
- Trabucchi Pietro
- Van Dijk Hans
- Van Megen Ron
- Vedana Fabio
- Venuto Tom
- Vicini Marcello
- Vino Giuseppe
- Volf Nadia
- Vorderman Carol
- Vroemen Guido
- Waterbury Chad
- Waters Paul
- Williamson Lexie
- Wissel Hal
- Young Megan
- Zanon Daniela
Tipi di contrazione muscolare nel fitness

Contrazioni dinamiche e contrazioni statiche o isometriche
Anche se spesso, parlando di fitness, si parla dei diversi tipi di contrazione muscolare, in realtà bisogna precisare che da un punto di vista puramente biomeccanico le definizioni e le divisioni sono spesso artificiose, inesatte o al meglio semplificate. Essenzialmente possiamo distinguere tra contrazioni dinamiche e contrazioni statiche o isometriche. In questo estratto dal libro Principi di Metodologia del Fitness di Antonio Paoli, Marco Neri e Antonino Bianco, vediamo insieme:
- Le contrazioni dinamiche ed isometriche;
- La contrazione isocinetica.

Le contrazioni dinamiche ed isometriche
Una contrazione si dice isometrica quando il muscolo viene stimolato a lunghezza costante, senza che i suoi estremi si avvicinino. In laboratorio questa è la tecnica di elezione per determinare la forza di un muscolo in funzione del tempo. Si esegue fissando gli estremi del muscolo a due punti fissi; in questo modo essi non possono avvicinarsi, ma comunque all’interno del muscolo la materia contrattile si accorcia, allungando i tendini e altri elementi elastici. Mentre il muscolo si accorcia, la materia contrattile compie un lavoro meccanico interno positivo, che va ad aumentare l’energia potenziale elastica.
Vi è contrazione isometrica in due occasioni:
- Quando si deve contrastare una forza esterna (gravità oaltro), ad esempio nel sostenere un peso a braccia tese oppure nel tentativo di vincere la resistenza di un oggetto troppo pesante;
- Quando c’è una co-contrazione di muscoli agonisti e antagonisti che porta a un’immobilizzazione del segmento scheletrico: sono tutte le contrazioni muscolari statiche che noi eseguiamo volontariamente senza carichi esterni, ad esempio le pose culturistiche.
La contrazione dinamica rappresenta un argomento più complesso e spesso, come già accennato, fonte di confusione anche per gli stessi autori. Diciamo subito che le contrazioni dinamiche che avvengono durante la normale vita di reazione si dividono in concentriche ed eccentriche, spesso unite da alcuni autori sotto la denominazione di contrazioni isotoniche, denominazione purtroppo invalsa nell’uso corrente ma errata. Infatti, “la contrazione isotonica è la contrazione durante la quale la tensione muscolare rimane costante mentre il muscolo si accorcia” (Wells, 1990). Risolutiva a questo punto è la precisazione di McArdle, Katch e Katch (1996) che affermano:
“Le contrazioni concentrica ed eccentrica sono comunemente denominate isotoniche perché in entrambi i casi c’è movimento. Il termine isotonico deriva dal greco “stessa tensione” (in inglese usano i termini strain e tension). In realtà questo termine è impreciso quando applicato alla maggior parte delle azioni muscolari dinamiche che comportano movimento, perché la capacità effettiva di generare forza varia con il variare degli angoli articolari; quindi la massima espressione di forza non è uniforme durante il ROM”.
NOTA
Il muscolo si accorcia durante la contrazione isometrica?
Durante una contrazione isometrica si accorciano sia i tendini (ad esempio, il tricipite surale circa dell’8-10%; Hof et al.,1998) sia le fibre. L’unione delle caratteristiche elastiche delle parti all’interno delle fibre (code della miosina) e dei tendini sono stati chiamati da Hill “elementi elastici non smorzati” perché sono capaci di accorciarsi molto rapidamente senza essere ostacolati da rilevanti forze viscose (Cavagna, 1988). Gli elementi costituiti dalle code miosiniche sono chiamati elementi elastici in parallelo, mentre i tendini e le aponeurosi elementi elastici in serie. La sola materia contrattile durante una contrazione isometrica si accorcerebbe dello 0.4% (Cavagna e Citterio, 1974). In realtà, ramificazioni tendinee si infiltrano nel muscolo, quindi, anche escludendo i tendini esterni, l’accorciamento elastico del muscolo non si identifica con l’accorciamento elastico della materia contrattile, perché le parti tendinee interdigitali con il muscolo ne influenzano le caratteristiche. Così, grazie alla partecipazione degli elementi elastici in serie, il sartorio di rana si accorcia del 2% della lunghezza dell’intero muscolo (rispetto all’ipotetico 0.4% imputabile alla sola materia contrattile), del 4-5% il gastrocnemio. Chiaramente, maggiore è la parte tendinea maggiore sarà la possibilità di accorciamento.
L’equivoco è nato dalla trasposizione sic et simpliciter dei lavori di laboratorio sul muscolo isolato e il muscolo in vivo, senza le dovute conoscenze di biomeccanica e fisiologia muscolare (Enoka, 2001; Wells, 1998). Suggeriamo a questo punto di chiamare le contrazioni eccentriche e concentriche in vivo anisotoniche. Il prof. Cavagna illustra ottimamente come in laboratorio si possa lavorare in condizioni di isotonia. Infatti, in vitro per mezzo della leva isotonica si ottiene di fare accorciare il muscolo contro un carico “effettivo” praticamente costante: ciò permette di evitare l’accorciamento dei tendini e quindi di fare coincidere l’accorciamento muscolare con quello della sola materia contrattile.
Chiarito questo punto scottante, possiamo soffermarci sulle altre contrazioni dinamiche da suddividere in concentriche ed eccentriche. La contrazione concentrica è quella che porta effettivamente all’accorciamento muscolare, con conseguente avvicinamento dei capi articolari. Nella contrazione eccentrica, invece, il muscolo funge da freno all’azione, contraendosi comunque ma venendo “forzato” a distendersi e tornando alla sua lunghezza di riposo. Questa è una capacità eccezionale del motore muscolare umano che lo differenzia in maniera significativa dagli altri motori, infatti anche durante questo lavoro negativo la forza del muscolo è diretta verso il centro del muscolo come nel lavoro positivo. È importante ricordare come il diagramma forza-velocità (studiato su contrazioni isotoniche) sia valido anche in vivo e ci indichi la funzione motrice del muscolo, descritta come un’iperbole a traslazione di assi dall’equazione di Hill:

La velocità massima si raggiunge con carico uguale a zero ed è impossibile da raggiungere perimentalmente poiché il peso della leva isotonica è comunque superiore a zero.

Questa è la velocità intrinseca del muscolo, cioè la rapidità dell’attività ATPasica miosinica.
È interessante sottolineare che con valori di forza superiori all’isometrico e velocità negative, corrispondenti alla fase eccentrica, per piccoli valori di velocità di allungamento abbiamo un brusco aumento della forza fino al 50% in più rispetto alla forza isometrica, che poi si mantiene costante.
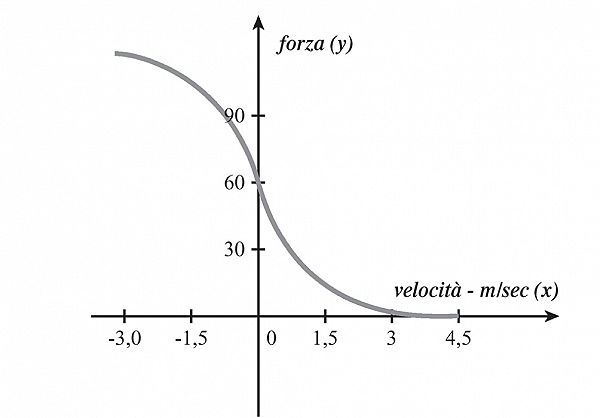
Fig. Rappresentazione schematica del grafico forza-velocità di Hill.
La contrazione isocinetica
Un altro tipo di contrazione dinamica è quella isocinetica, che è eseguibile solamente con particolari apparecchiature. Nata anch’essa in laboratorio, è stata poi trasportata in vivo con speciali macchine che permettono una contrazione a velocità costante, grazie a particolari sistemi di feedback che regolano automaticamente lo sforzo imposto al segmento articolare. Altre suddivisioni del tipo di contrazione, utilizzata per lo più in ambito sportivo ma molto poco in fisiologia muscolare, sono:
- Contrazioni auxotoniche: dove, con l’accorciarsi del muscolo, aumenta la tensione del carico esterno (elastici);
- Contrazioni pliometriche: dove a un rapido allungamento.
Segue immediatamente una contrazione per sfruttare l’energia elastica accumulata negli elementi elastici.



